
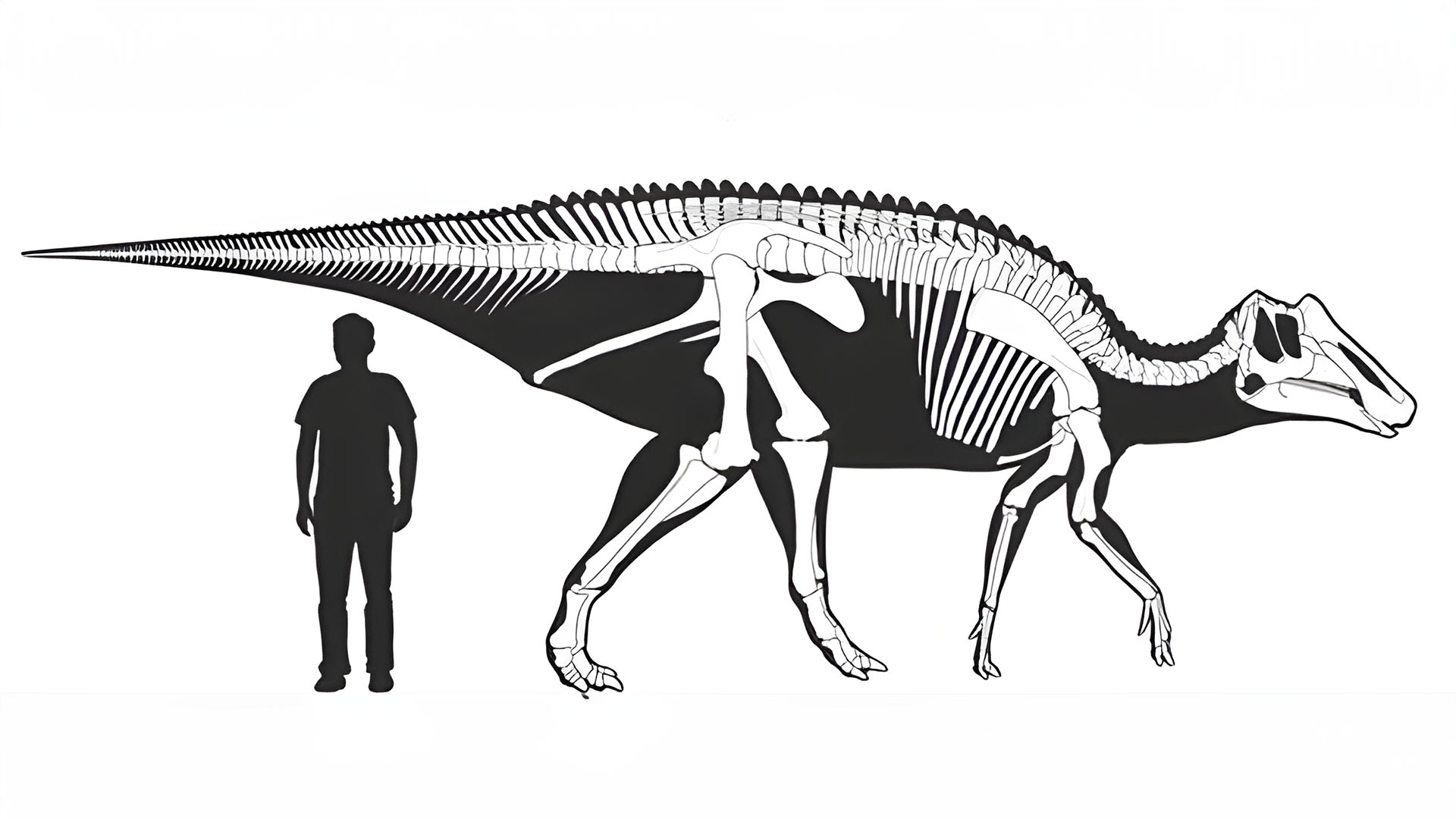
Image Credit: McGarrity, C. T., Campione N. E. , and Evans D. C. (2013) “Cranial anatomy and variation in Prosaurolophus maximus (Dinosauria: Hadrosauridae)”.
John Michalski is a senior geology major at the University of Wisconsin, Milwaukee. He has been passionate about dinosaurs since he could walk and talk, and loves to discuss and educate others about paleontology and prehistoric life.
Although it lacks the extraordinary headgear, famous dig sites, or even the novelty of being Tyrannosaurus rex’s lunch like its relatives, Prosaurolophus remains an important piece in the puzzle of understanding the world (specifically western North America) during the last several million years of the Mesozoic. On a personal note, I did a video project about Campanian dinosaurs a while back, and Prosaurolophus was a central focus of the video. As such I have a bit of a fondness for it, and I see no reason why this guy can’t get the same amount of love as other, more well-known hadrosaurs.
- Prosaurolophus, not to be confused with the somewhat more famous Saurolophus, was a fairly large saurolophine hadrosaur from the late Cretaceous of North America. Though known material does not describe an immense animal, current estimates place its length at around 8.5 meters. Its fossils have been dated to the latter half of the Campanian stage (about 75 million years ago). Remains of this dinosaur have been found in Alberta, Canada, as well as Montana within the United States.
- Prosaurolophus shared its domain with many other kinds of dinosaurs. In Alberta, fellow herbivores Styracosaurus and Lambeosaurus foraged alongside it. In Montana, Einiosaurus, Scolosaurus, and Hypacrosaurus were contemporary plant-eaters. In both areas, Prosaurolophus may have been preyed upon by the fearsome tyrannosaur, Daspletosaurus.
- There are two known species of this dinosaur: Prosaurolophus maximus, the type species, was found in 1915 and described a year later by Barnum Brown. Its bones came from Dinosaur Provincial Park in southern Alberta, Canada, specifically from the later Campanian of the Dinosaur Park formation. A second species, Prosaurolophus blackfeetensis, was described by Jack Horner in 1992, from the younger sediments of the upper Two Medicine Formation in Montana. Subtle differences in the small crest on top of their skulls, according to Horner, help distinguish them from one another. Other scientists have suggested, however, that P. blackfeetensis is the same animal as P. maximus.
- A bonebed of Prosaurolophus from the Two Medicine Formation suggests that these dinosaurs may have been herding animals, a behavior often depicted among other hadrosaurs.
- Some scientists have interpreted bony ridges around the nostril area as mounting points for a fleshy air sac that may have served as a display structure by males to attract mates, as well as amplify calls to enhance communication amongst individuals.
- Prosaurolophus was part of a study analyzing the scleral ring (ring of bones that supports the eye) and orbit (eye socket) of thirty-three different archosaurs. Though the eyeballs themselves decay away before any other part of the body, scleral ring bones do sometimes fossilize. Some scientists claim that the shapes of scleral bones can indicate the shape of the eye; this idea remains somewhat controversial, as others point out that thin bones like these can change shape during the process of fossilization. However, if properly managed, the data from the eyes of these sampled species might help determine when they would have been up and about throughout a twenty-four-hour period. With Prosaurolophus, the study indicated that it may have been cathemeral, meaning that it was active sporadically during the day.
- Prosaurolophus’ distribution from Canada to Montana may have allowed it to adapt to different ecosystems. The Dinosaur Park Formation has been interpreted as once being a warm, lush landscape made up of rivers and flood plains. Meanwhile the Two Medicine Formation has been interpreted as being drier, and prone to seasonal droughts. It’s possible that mortality rates may have risen during these leaner times, which could be a factor in explaining the numerous bonebeds found at the formation. However, based on its finds in two very different habitats, Prosaurolophus may have been able to cope with the harsher climate. It has been suggested, though far from confirmed, that this shift in climate encouraged Prosaurolophus to expand its range, perhaps evolving into the Maastrichtian epoch’s Saurolophus and Augustynolophus.
- Interestingly, the environmental shift in Prosaurolophus’ world is strikingly similar to the shift in habitat of the large ornithopod Tenontosaurus of the early Cretaceous, if reversed. When it first appeared in North America, Tenontosaurus’ world was arid and dry, but became increasingly wet and tropical as millions of years passed. This was likely due to the birth of the Western Interior Seaway which divided North America into two landmasses during Prosaurolophus’ time. In much the same way as suggested for Prosaurolophus, this change allowed for Tenontosaurus to expand in its range, indicating that it too was a very adaptable dinosaur.
- The ecological roles hadrosaurs played in their environments is not well understood, as their diets are variable. Fortunately, one Prosaurolophus jawbone from Alberta gives us an idea as to what it may have had to satisfy its appetite. The jaw bears an unusually large piece of amber attached to the bone. Amber is fossilized tree resin (it’s the stuff Jurassic Park used to get DNA for its dinosaurs), and unlike tree sap, it is poisonous. It seems a bit weird, and highly unlikely, that a hadrosaur would be eating resin for all of its life, unless it decided it was better off poisoning itself and saving it the trouble of being chased down by a hungry tyrannosaur.
- But then scientists found another clue within the amber itself: a type of aphid, specifically one part of a family that primarily targeted conifer trees. As luck would have it, the amber itself belonged to a tree from the genus Cupressus, a type of evergreen conifer. As resin is intended as a defense mechanism in response to injuries made upon the tree by insects or other animals, the aphid may have been caught by the tree’s defenses. Based on what the amber and the aphid show us, Prosaurolophus was likely a browser, feeding from tall trees rather than shrubs or ferns.
- As for how the resin ended up in the hadrosaur’s mouth, it’s perhaps the subject of another study. It could’ve been an odd coincidence, but then again . . . who’s to say something far stranger, maybe even darker is at play here? Could it have been Ms. Scarletosaurus, in the Campanian of Alberta, with the resinous goo? The world may never know.
Sources:
Christopher T. McGarrity, Nicolas E. Campione, David C. Evans, Cranial anatomy and variation in Prosaurolophus maximus (Dinosauria: Hadrosauridae), Zoological Journal of the Linnean Society, Volume 167, Issue 4, April 2013, Pages 531–568, https://doi.org/10.1111/zoj.12009
Eberth, David A. “3. The Geology.” Dinosaur Provincial Park: a spectacular ancient ecosystem revealed (2005): 54.
Forster, Catherine A. “The paleoecology of the ornithopod dinosaur Tenontosaurus tilletti from the Cloverly Formation, Big Horn Basin of Wyoming and Montana.” The Mosasaur 2 (1984): 151-163.
Hopson, J. (1975). The evolution of cranial display structures in hadrosaurian dinosaurs. Paleobiology, 1(1), 21-43. doi:10.1017/S0094837300002165
Lull, Richard Swann; Wright, Nelda E. (1942). Hadrosaurian Dinosaurs of North America. Geological Society of America Special Paper 40. Geological Society of America. p. 226.
McKellar, R.C., Jones, E., Engel, M.S. et al. A direct association between amber and dinosaur remains provides paleoecological insights. Sci Rep 9, 17916 (2019). https://doi.org/10.1038/s41598-019-54400-x
Rogers, Raymond R. “Taphonomy of Three Dinosaur Bone Beds in the Upper Cretaceous Two Medicine Formation of Northwestern Montana: Evidence for Drought-Related Mortality.” PALAIOS, vol. 5, no. 5, 1990, pp. 394–413. JSTOR, www.jstor.org/stable/3514834. Accessed 4 Jan. 2021.
Schmitz L, Motani R. Nocturnality in dinosaurs inferred from scleral ring and orbit morphology. Science. 2011 May 6;332(6030):705-8. doi: 10.1126/science.1200043. Epub 2011 Apr 14. PMID: 21493820.